mean annual precipitation
and temperature (e.g. Weimann et al 1999), have not been widely investigated
such that absences of certain characters or character states in these fossils may
simply be due to the fact that they have not yet evolved! Thirdly, these
formulae rely on large sample sizes, larger than we have yet studied. However,
we are increasing our sample size of fossil material such that we will be able
to understand more fully the state of wood anatomical character evolution in
the Southern Hemisphere and thus be able to provide more rigorous climatic
indications and enhance the validity of such a taxonomic independent approach.
|

Antarctica 60 million
years ago?
Similar
vegetation is found today in temperate forests of Chile and Southern
Australia (photo taken whilst on fieldwork in southern Australia)
|
Another important focus
is that of biodiversity change from the Cretaceous to Tertiary. High
latitude Cretaceous floras differ wildly from those at mid-latitudes. High
latitude floras contained a diversity and abundance of certain plant groups
that are either absent or ecologically unimportant in lower latitudes. One
such example is the liverwort- and moss component. High latitude floras such
as those of the Antarctic Peninsula or southeastern Australia contain up to
20% within-floral diversity whereas in mid-latitude floras they are rare or
absent. Furthermore, certain groups seem to survive for longer at high
latitudes than lower latitudes. The bennettites and other types of gymnosperm
persist into the Late
|
Cretaceous (as recent as
Santonian), yet are probably extinct from low latitudes by Albian times. Why
plant groups such as these are more suited to extreme climatic conditions of
warm polar latitudes is still unknown although the difference in composition is
largely tied to one of the major biotic cataclysms in Earth History: The
origin, radiation and diversification of flowering plants which had profound
effects on the vegetation. Araucarian- (monkey puzzle trees) and podocarp
conifers dominated the pre-angiosperm overstorey with rarer components
including Ginkgo (maiden hair tree).
The understorey contained abundant and diverse fern and bennettite communities.
The angiosperms initially expanded into the understorey replacing the
bennettites and ferns. During the ensuing 40 million years or so, up until the
early Tertiary, angiosperms increased their advance to appropriate the mid- and
overstorey niches. Ferns were able to make a comeback through exploiting new
habitats created by angiosperms and consequently recovered in numbers. The
conifers however were pushed further into increasingly unfavourable conditions
resulting in their ultimate decline in numbers (Cantrill and Poole in press).
Angiosperm
diversification brought colour, taste and smell to the Antarctic
landscape through their flowers, seeds, fruits, leaves and bark. Evidence from
the macrofossils record gives us a taste of what Gondwana might have been like:
The presence of, amongst others, the Illiciaceae (or star anise) with flowers
of red or yellow smelling of fish but with aniseed
flavoured seeds and aromatic leaves used extensively in authentic eastern cuisine
today. Bundles of conspicuous white and cream flowers, and seeds and
wood tasting and smelling of pepper would have been indicative of the
presence of the Winteraceae. The avocado pear family is represented by Sassafras with its pendulous bright yellow
flowers set against pale green foliage that exudes an aroma reminiscent of
fresh lemons and later turn to autumnal shades of orange and red.
Members of the Monimiaceae were present with their edible fruits, and leaves
and bark that produce aromatic oils, commercially exploited by
the international perfume industry today. Canopies with breathtaking
hues of yellow, red and orange would be accredited to the abundant Nothofagus (southern beech), a taxon
today only found across austral landmasses but cultivated globally for their
autumnal foliage.
Sadly, it should be noted that these
fragile, primeval Valdivian forests, the only surviving representative of
Antarctica’s Late Cretaceous-Early Tertiary vegetation, are being felled
uncontrollably. The passing away of these forests will take with it the chance
to further our understanding of the ecological diversity and dynamics. With
that, an important legacy of a unique ecosystem that once existed some 80 - 40
million years ago in the southern high latitudes of Gondwana, will be lost
forever.
Funding: This work was initiated as a NERC funded project but
continues with the close collaboration from colleagues in the UK especially Dr
David Cantrill at the British Antarctic Survey, U.K.
|
B. Lower Tertiary paratropical pyritised wood flora from southern England
|

A desolate beach in SE England today was
once a subtropical paradise
|
Results from this work so far
The
Eocene London Clay flora from southern England is a classic example of a boreotropical
flora and represents one of the most varied, and the only diverse, flora from
lower Eocene strata in Europe. The London Clay flora, with approximately 145
living and extinct genera placed in living families, shares similarities with
smaller floras from this period in Europe and North America and suggests that a
belt of similar vegetation may have extended across the Northern Hemisphere
during the lower and early middle Eocene (Collinson, 1983).
|

|
The wood forms a major
constituent of this flora but has been studied comparatively little but the
wood component reveal taxa, for example the Dipterocarpaceae, as yet
unknown in the fruit and seed flora. Continued studies focusing on the wood
component will help in understanding the composition of this boreotropical
vegetation. Identifications of the wood flora include the Vitaceae,
Sapindaceae, Platanaceae, Celastraceae, Pinaceae, Taxodiaceae and at least 6
fern types of which one is a polypodiaceous epiphyte.
|
|
Pyritised wood from the Southeast
of England belonging to the Platanaceae (plane tree family)
|
|
All
the records provide significant information concerning the diversity of the
vegetation that covered southern England during the lower Eocene. Using the Nearest
Living Relative approach, evidence from the wood flora supports a postulated paratropical
environment similar to that of Indo-Malaysia today. Seasonality has been
suggested from the presence of indistinct, or even absence, of growth rings in
the wood which may be indicative of a more tropical, rather than a temperate,
flora. However, growth ring data need careful analysis as growth rings
can vary according to genera rather than climate (Détienne, 1989). Elements
found in the wood flora also serve to illustrate that certain taxa, such as the
Cercidiphyllaceae, were once widespread across the Northern Hemisphere in the
geological past (Crane 1984). Inclusion of the taxonomic data from fossil wood,
including studies of plant animal interactions as evidenced from frass and
borings in the London Clay wood flora will contribute to a greater
understanding of the biodiversity and prevailing climate in southern
England and increase our knowledge concerning Lower Eocene boreotropical
floras.
Funding: This work was initiated through NERC funding and
continues through close collaboration with colleagues from the Jodrell
Laboratory, Royal Botanic Gardens, Kew, U.K.
|
C. Late Tertiary tropical
wood from the Amazon Basin, Peru
|

|
Results
from this work so far:
Field work in 2001 enabled
collections of mummified wood predominantly from
fossil log jams (palisadas). Mummified tree trunks up to 2 m in diameter were
excavated from these fossil log jams in sections dating back to the Miocene within
the Manu National Park. Material of this age can be identified to genus level.
|

|

|
|
Miles Silman (leaft) and Pim van Bergen (right)
standing by Miocene fossil tree trunks, many metres in length, being washed
out of the river bank
|
These woods record
details of past environments in both their anatomical structure and geochemical
signatures. Palaeoecological reconstruction based on
palynological evidence is very successful at demonstrating both dynamics and
composition of individual tree communities in temperate floras. However, tropical
forest communities are nearly an order of magnitude more diverse. Problems
involving the non-appearance of important indicator species in the pollen
spectra coupled with the fact that most tropical species are entomophilous
(insect pollinated and therefore do not produce copious amounts of pollen),
indicates that a combined palynological-macrofossil (e.g. wood) approach will
provide the most representative data on community structure and dynamics.
Coupling this data with evidence from the (more spatially biased) pollen flora
and species diversity lists of the modern flora, vegetation composition and
dynamics within the Rio Manu floodplain can be assesed in response to changing
climate from Miocene to present.
Taphonomic studies were undertaken on the modern
log jams within the Manu River to (1) qualify the bias today which can then be
compared with possible biases that might be found in the fossil material, and
(2) determine the taxonomic composition and thus whether log jam material
reflects the current vegetation along the Manu River. This data can then be
used when interpreting and drawing conclusions from the fossil wood assemblages
(also found in log jams). It was found that the wood content of the modern log
jams are biased in favour of successional forest and does not represent the
vegetation in the Manu reserve. The propagule assemblage however reflected a
more regional bias. Only together could the natural vegetation be
reflected in the dispersed assemblage and even then there were biases with
regard to organ and crop size and density.
Funding:
Grant from the Percy Sladen Memorial
Fund and is being undertaken in close
collaboration with Dr Miles Silman from Wake Forest University, North Carolina,
U.S.A.
2. Molecular
palaeoclimatic interpretations from fossil plant remains
Results
from this work so far: To date, stable carbon isotope
analyses of marine carbonates and bulk organic carbon are often used as a basis
for palaeoclimate reconstructions. In recent years the use of individual
organic molecules has been gathering momentum allowing more precise
distinctions between different sources contribution to the d13C
signals. However, most of this research has focused on marine sourced organic
matter or on molecules the origin of which is less unambiguous (i.e. alkanes).
My current project is focusing on terrestrially derived fossil and modern wood
using compound specific stable carbon isotope analyses to evaluate
changes in past and palaeo- atmospheric CO2 concentrations.
Combining molecular and bulk data for young (i.e. post Miocene) material
chemical taphonomy (in terms of selective loss of certain moieties, e.g.
polysaccharide or lignin) affect the bulk values. For older fossils internal
chemical transformations cause additional isotopic shifts that are more
ambiguous and are currently being studied.
Funding: This work is being financed by NWO/AWL and is
undertaken in collaboration with colleagues in the Netherlands
Institute for Sea Research (NIOZ) and at RING.
3. Natural variation in
modern systems – understanding the signals locked within fossils
Acknowledging
variation raises problems with regard to data interpretation, it hinders
attempts to incorporate data from living organisms into computer generated
models and above all it requires more interpretation (and imaginative?)
deliberation to explain the data at hand. Variation is an integral part of
organismal characteristics and has important implications for ecology,
palaeoecology and evolutionary theory. Therefore maybe variation can be used to
our scientific long term advantage and should now be recognised and discussed
rather than simply ignored. My research takes into account variation and looks
to determine the extent of variation such that the data obtained can be
meaningful and conclusions regarding ecological systems through the geological
past can be substantiated. This focus is incorporated into the above projects.
An example of variation follows:
|
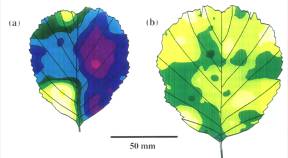
|
Variation in stomatal density in sun (a)
and shade (b) leaves of Alnus glutinosa. Similar ranges
can be seen with stomatal index. Range from light yellow with 100-120 stomata
per mm2 to light purple >240 stomata per mm2 (Poole
et al. 1996).
|
Publications
Poole I. (2002)
Systematics
of Cretaceous and Tertiary Nothofagoxylon: Implications for Southern
Hemisphere biogeography and evolution of the Nothofagaceae. Australian
Systematic Botany (in press).
van Bergen P.F. and Poole I. (2002)
Stable
carbon isotopes in wood: A clue to palaeoclimate? Palaeogeography, Palaeoclimatology, Palaeoecology (in press).
Cantrill D.J. and Poole I.
(2002)
Cretaceous
patterns of floristic change in the Antarctic Peninsula. Geological Society
of London Special Publication (in press).
Francis J.E.
and Poole I. (2002)
Cretaceous
and Tertiary climates of Antarctica: evidence from fossil wood. Palaeogeography, Palaeoclimatology,
Palaeoecology (in press).
Poole I. and Cantrill D.J. (2001)
Forest of the polar night. Geoscientist 11(9), 4-6.
Poole I. and Cantrill D.J. (2001)
Fossil woods from
Williams Point Beds, Livingston Island, Antarctica: a Late Cretaceous southern
high latitude flora. Palaeontology 44,1081-1112.
Poole
I. and Davies C. (2001)
Glutoxylon Chowdhury (Anacardiaceae): the first known
record of fossil wood from Bangladesh. Review
of Palaeobotany and Palynology 113, 261-272.
Poole
I. and Gottwald H. (2001)
Monimiaceae sensu lato, an element of Gondwanan polar forests: Evidence from the Late
Cretaceous-early Tertiary wood flora of Antarctica. Australian Systematic Botany 14, 207-230.
Poole I.. Hunt R.J. and Cantrill D.J.
(2001)
A
fossil wood flora from King George Island: ecological implications for an
Antarctic Eocene vegetation. Annals of
Botany 88, 33-54.
van Bergen P.F. and Poole I. (2001)
Biasing the stable isotope record: The effect of
chemical taphonomy on fossil plant remains. Terra Nostra 2001/3,
219-222.
van
Bergen P.F. Poole I. Oglivey T. Capel C. and
Evershed R.P. (2000)
Evidence for
demethylation of syringyl moieties in archaeological wood using Pyrolysis/Gas
Chromatography/Mass Spectrometry. Rapid
Communications in Mass Spectrometry 14, 71-79.
Poole I. (2000)
Variation - Nature’s
spanner or an unrecognised tool? Palaios
Online 15, 1-2.
Poole
I. (2000)
Fossil angiosperm wood
anatomy: its role in the reconstruction of biodiversity and palaeoenvironment. Botanical Journal of the Linnean Society
134, 361-381.
Poole I.. Cantrill D.J., Hayes P. and
Francis J.E. (2000)
The
fossil record of Cunoniaceae: new evidence from Late Cretaceous wood of
Antarctica. Review of Palaeobotany and
Palynology 111, 127-144.
Poole
I. and Francis J.E. (2000)
The first record of
fossil wood of Winteraceae from the Upper Cretaceous of Antarctica. Annals of Botany 85, 307-315.
Poole
I., Gottwald H. and Francis J.E. (2000)
Illiciaceae, an element of Gondwanan polar forests?
Late Cretaceous and Early Tertiary woods of Antarctica. Annals of Botany 86, 421-432.
Poole
I., Lawson T., Weyers J.D.B. and Raven J.A.
(2000)
Effect of elevated CO2
on the stomatal distribution and leaf physiology of Alnus glutinosa (L.) Gaertn.. New
Phytologist 145, 511-521.
Poole
I. and Lloyd, G.E. (2000)
A new SEM technique for
observing pyritised plant material. Review
of Palaeobotany and Palynology 112, 287-295.
Poole I. and Page C. (2000)
A fossil fern indicator
of epiphytism in a Tertiary flora. New
Phytologist. 148, 117-125.
Poole
I.. Richter H. and Francis J.E. (2000)
Gondwanan origins for Sassafras (Lauraceae): evidence from
Late Cretaceous fossil wood of Antarctica. International
Association of Wood Anatomists Journal 21, 463-475.
Poole I. and Wilkinson
H.P. (2000)
Early
Eocene vines of Southeast England. Botanical
Journal of the Linnean Society 133, 1-26.
Poole I. (1999)
The
presence and absence of growth ring structures in fossil twig wood: some
possible explanations. In The Evolution
of Plant Architecture. Eds A.R. Hemsley & M. Kurman. Royal Botanic
Gardens, Kew, pp. 205-219.
Poole I. and Francis
J.E. (1999)
The
first record of fossil atherospermataceous wood from the upper Cretaceous of
Antarctica. Review of Palaeobotany and
Palynology 107, 97-107.
Poole
I. and Francis J.E. (1999)
Reconstruction of
Antarctic palaeoclimates using angiosperm wood anatomy. In: Proceedings of the 5 EPPC, Acta
Palaeobotanica supplement 2 173-179.
Poole I. and Kürschner W. (1999)
Stomatal
density and index: the practice. In: Fossil
Plants and Spores: modern techniques
Eds Jones T.P. & Rowe N.P.
Geological Society London, pp. 257-260.
Poole I. and Wilkinson
H.P. (1999)
A
celastraceous twig from the London Clay (Eocene) of Southeast England. Botanical Journal of the Linnean Society
129, 165-176.
Lockheart M., Poole I., van Bergen P.F. and
Evershed R. (1998)
Leaf
carbon isotope compositions and stomatal characters: important considerations
for palaeoclimate reconstructions. Organic
Geochemistry 29, 1003-1008.
Poole I. (1996)
Conifer
twigs from the London Clay (Eocene) of Southeast England. Review of Palaeobotany and Palynology 94, 25-37.
Poole I. Weyers J.D.B. Lawson T. and
Raven J.A. (1996)
Variations
in stomatal density and index: implications for palaeoclimatic reconstructions.
Plant, Cell and Environment 19,
705-712.
Poole I. (1994)
Twig
wood anatomical characters as palaeoecological indicators. Review of Palaeobotany and Palynology 81, 33-52.
Poole I. (1993)
A
dipterocarpaceous twig from the Eocene London Clay Formation of Southeast
England. Special Papers in Palaeontology
49, 155-163.
Poole I. (1992)
Pyritized
twigs from the London Clay, Eocene, of Great Britain. Tertiary Research 13, 71-85.
Poole I. and Wilkinson
H.P. (1992)
Two
sapindaceous woods from the London Clay (Eocene) of Southeast England. Review of Palaeobotany and Palynology
75, 65-75.